15 June 2022: Animal Study
Differential circRNA and mRNA Expression Profiling During Osteogenic Differentiation in Rat Bone Marrow Mesenchymal Stem Cells
Yuan Yao1ABCDEFG, Jialin Liu1BDE, Wenjing Zhang2BC, Xiangzhen Han1G, Huiyu He1AD*, Xiaowei Ji1DEDOI: 10.12659/MSM.936761
Med Sci Monit 2022; 28:e936761






Abstract
BACKGROUND: Circular RNAs (circRNAs) play important roles in gene expression and signaling pathways. The study aimed to identify the differential expression of circRNAs and mRNAs in the osteogenic differentiation of rat bone marrow-derived mesenchymal stem cells (rBMSCs) and to explore the biological function of circRNAs in the osteogenic differentiation of rBMSCs.
MATERIAL AND METHODS: High-throughput sequencing was used to detect differentially expressed circular RNA and mRNA during osteogenic differentiation of rBMSCs. The RNAs were analyzed for Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment to predict their potential role in regulating rBMSC osteogenesis. MiRanda, Circatlas, and miRDB databases were used to predict target relationships between circRNA, miRNA, and mRNA.The regulatory network was constructed by Cytoscape (version 3.6.1). The RNA-Seq findings were validated by quantitative real-time PCR (qRT-PCR).
RESULTS: The results revealed that 29 differentially expressed circRNAs and 2453 differentially expressed mRNAs were detected during the osteogenic differentiation of rBMSCs. Many differentially expressed circRNAs were closely related to osteogenic differentiation of cells. Among them, circRNAs_1809 and Kitlg were the significantly increased circRNA and mRNA during osteogenic differentiation of rBMSCs. The ceRNA network showed that circRNA_1809 could target the Kitlg gene through miR-370-3p.
CONCLUSIONS: CircRNAs may play an important role in the osteogenic differentiation of rBMSCs. CircRNA_1809 may acts as a sponge for miR-370-3p and regulate the osteogenic differentiation of rBMSCs by targeting Kitlg; however, this hypothesis needs further verification. This study laid a theoretical foundation for further understanding the mechanism of circRNAs regulating osteogenic differentiation of rBMSCs.
Keywords: Mesenchymal Stem Cells, Osteogenesis, RNA, Circular, Animals, Gene Expression Profiling, MicroRNAs, RNA, Messenger, Rats
Background
Currently, autologous bone transplantation remains the criterion standard for treating bone defects. However, not all kinds of bone xenograft materials can wholly repair bone tissue structure and function. Therefore, it is crucial to develop bioactive materials to regulate the osteogenic differentiation of cells [1]. As seed cells, rat bone marrow-derived mesenchymal stem cells (rBMSCs) can self-renew and multi-direct differentiation. They can also differentiate into osteoblasts, adipocytes, chondrocytes, muscle cells, and even nerve cells. Moreso, the cells have been found to possess less immune rejection, easy access, and value-added ability. As a result, they have become an ideal cell for bone tissue engineering [2–5].
Circular RNAs (circRNAs) are different from general linear RNAs due to the lack of a 5’ cap structure and 3’ polyadenylation tail that form a unique closed-loop structure with covalent bonds. Compared with linear RNA, they are not easily degraded by exonuclease RNase R and possess a more stable structure [6]. They are widely distributed in eukaryotes and regulate gene expression [7]. It is proven that circRNAs are involved in embryonic development, cell activities, and many human diseases. Therefore, ongoing research of circRNAs has become a new hotspot in the field of RNA [8]. Furthermore, there is evidence that circRNA have microRNA (miRNA) binding sites and usually act as a sponge of miRNA to competitively bind miRNA in cells; the expression of miRNA in cells is reduced, thus weakening the inhibitory effect of miRNAs on their downstream mRNAs. The expression level of mRNA is indirectly increased, and this competitive way of regulating gene expression is called competitive endogenous RNA (ceRNA). Based on this principle, the ceRNA regulatory network composed of circRNA-miRNA-mRNA can be constructed by bioassay technology, providing a research direction for studying the mechanism of circRNA regulating gene expression [9]. For example, circSLC8A1 acts as a sponge for Mir-130b/Mir-494 and inhibits bladder cancer progression by regulating PTEN [10]. Although many experiments demonstrated that circRNAs play a crucial role in cancer and cardiovascular disease, a shortage of information exists regarding the relationship between circRNAs and osteogenic differentiation. Previous studies have detected the differential expression of circRNAs between rDFC and PDLSC during osteogenic differentiation by high-throughput sequencing technology [11,12]. Notwithstanding, the regulatory role of circRNAs in the osteogenic process of rBMSCs has not yet been established.
Although some studies have proven that circRNAs and miRNAs are essential regulators in cell osteogenesis, their potential interaction and regulatory mechanism remain unclear. Therefore, in this study, we used high-throughput sequencing technology to detect the circRNA expression profile in the process of osteogenic differentiation of rBMSCs. We also predicted that circRNAs would provide a regulatory mechanism for the osteogenic differentiation of rBMSCs through Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG). Furthermore, bioinformatics was used to analyze and construct the circRNA-miRNA-mRNA competitive regulation network. Finally, quantitative real-time PCR (qRT-PCR) validated the circRNAs and mRNAs with significant differences in sequencing results.
Material and Methods
CULTURE OF BMSCS:
The animal procedure was approved by the Research Ethics Committee of the First Affiliated Hospital of Xinjiang Medical University (IACUC20170706-04). First, Sprague-Dawley (SD) rats were sacrificed after neck removal and placed in 75% ethanol solution for disinfection for 15 min. Then, in a sterile environment, the hind limbs were excised, and the soft tissues of the tibia and femur were removed. Next, the femur and tibia of the rats were separated and washed twice with PBS solution. Both ends of the bone were cut to expose the bone marrow cavity and then repeatedly rinsed with a medium containing 15% fetal bovine serum. After filtering through the cell sieve, the bone marrow cavity was collected into a 15-mL centrifuge tube, and the centrifuge was set to 160×g at 4°C for 10 min. After the supernatant was discarded, the isolated rBMSCs were inoculated in 25-cm2 tissue culture bottles and cultured in a humid atmosphere of 37°C and 5% CO2. The medium was changed every 3 days. When the cells reached 80% fusion, the cells were digested with 0.25% trypsin at 37°C for 2 min. After that, trypsin action was terminated by adding 10% culture medium with 3 times the volume of trypsin solution. The cells were tapped gently, then transferred into a 15-mL centrifuge tube and centrifuged for 5 min. Finally, the cells were divided into culture flasks according to the ratio of 1: 2 to continue the culture and complete cell passage.
FLOW CYTOMETRY IDENTIFICATION OF BMSCS:
The second-generation rBMSCs were washed twice with PBS, digested with 0.25% trypsin, and collected into three 1.5-mL EP tubes with 106 rBMSCs in each tube. Next, 10 μL containing 3% PBS was added into the EP tubes and allowed to stand for 30 min at 4°C. Then, under the condition of avoiding light, 2 μL of CD29 and CD452 antibodies were added to 2 EP tubes, respectively. Simultaneously, 2 μL PBS was added into EP tubes of the negative control group. The cells were incubated at 4°C for 30 min, washed with 1 mL pre-cooled PBS, and centrifuged at 160×g at 4°C for 5 min. After that, the remaining antibody and supernatant were discarded. Meanwhile, 500 μL PBS was added, homogenized, and placed in a flow cytometer for detection. Finally, after identification by flow cytometry, the third generation rBMSCs were selected for the subsequent experiments.
IMMUNOFLUORESCENCE IDENTIFICATION OF BMSCS:
First, the culture plate was washed thrice with PBS, the slides containing cells were fixed at 4% paraformaldehyde for 15 min and soaked with 0.5% Triton X-100 (PBS) at room temperature for 20 min. Next, the slides were washed with PBS 3 times and sucked dry with absorbent paper; then, normal goat serum was added to the slides and sealed at room temperature for 30 min. After that, the excess sealing fluid was sucked up with absorbent paper, and an aliquot of diluted primary antibody was added to the slides, later introduced into a wet box for incubation at 4°C overnight. Also, fluorescent secondary antibody was added in the dark after the climbing tablets with PBST were washed thrice. Next, the excess liquid on the climbing tablets was sucked up using absorbent paper, diluted fluorescent (Cy3) labeled Sheep anti-rabbit IgG was dropped, incubated in a wet box at 20 to 37°C for 1 h, and the slices were washed with PBST for 4 times. Finally, PBST was washed 4 times for 5 min to remove excess DAPI. Next, the liquid on the slipper was dried with absorbent paper. The slipper was then sealed with the slipper sealing solution containing an anti-fluorescence quenching agent. Lastly, the images were observed and collected under a fluorescence microscope.
OSTEOGENIC INDUCTION OF RBMSCS:
The third generation rBMSCs with good growth conditions were inoculated into 6-well plates at a density of 1×106 cells/well from the culture flask for further culture. When the cells reached 80% fusion, the medium was sucked out and washed twice with PBS. In the osteogenic induction group, 2 mL of osteogenic induction solution (10% fetal bovine serum+10 nmol/L β-sodium glycolate+100 nmol/L dexamethasone) was added into each well, and the control group was added ordinary medium. The culture medium was changed every 3 days, and cells were collected at the designated time interval.
ADIPOGENIC INDUCTION OF RBMSCS:
The third generation rBMSCs in the logarithmic growth phase to be induced were selected and inoculated into 6-well plates with a inoculation density of 1×106 cells/well. When the cell growth confluence reached 95% to 100%, the 10% complete medium was replaced with SD rat bone marrow mesenchymal stem cell adipogenic induction differentiation medium A (Cyagen Biosciences, China), and 3 days later, it was replaced with SD rat bone marrow mesenchymal stem cell adipogenic induction differentiation medium B (Cyagen Biosciences). Liquid A and liquid B were used alternately for 3 to 5 cycles until fat droplets formed.
ALIZARIN RED S STAINING AND QUANTIFICATION:
First, BMSCs were washed twice with PBS and fixed in 4% paraformaldehyde solution at 4°C for 30 min. Subsequently, the cells were washed with distilled water and stained with 0.1% alizarin red staining (Sigma Aldrich, USA) for 30 min. Next, after washing 3 times with distilled water, stained calcium nodules (Leica, Germany) were observed under a microscope. The dye was dissolved in 1 mL of 10% cetylpyridine (Sigma Aldrich) for 1 h, and the absorbance value was measured at 570 nm for quantitative analysis.
OIL RED O STAINING:
The oil red O raw solution was diluted with deionized water (3: 2) and centrifuged at 250×g to take the supernatant, which was the oil red O filtrate. The cells were washed with PBS twice and fixed in 4% tissue cell fixative solution at room temperature for 30 min. The fixative solution was discarded and washed with PBS twice. Staining was done with oil red O filtrate for 30 min, the staining solution was discarded, and the cells were washed with PBS twice. An amount of 1 mL PBS was added to each well, and the staining effect of fat droplets was observed under the microscope.
TOTAL RNA EXTRACTION OF RBMSCS:
Additionally, after 14 days of osteogenic induction or normal culture, 1 mL Trizol was added to each well of the 6-well plate, the bottom was blown repeatedly until the cells were fully lysed, and then they were transferred to the EP tube. Next, 200 μL chloroform was added and vortexed, and left to stand at 4°C for 5 min. It was then centrifuged at 4°C for 160×g for 10 min.
Next, the supernatant was transferred to a new EP tube, and 500 μL isopropyl alcohol was added slowly upside down and left to stand at room temperature for 30 min. Next, centrifugation was performed at 1600×g at 4°C for 10 min. Later, the supernatant was discarded, 500 μL pre-cooled 75% ethanol prepared with DEPC water was added, and the preparation was centrifuged at 1600×g, 4°C for 10 min. Again, the supernatant was discarded and allowed to dry at room temperature. Finally, 20 μL DEPC water was added to each tube to mix the RNA thoroughly. Lastly, the purity and concentration of RNA were measured using a spectrophotometer. The concentration of RNA solution was between 200 and 2000 ng/μL, and the purity was between 1.8 and 2.0 for subsequent experiments.
LIBRARY CONSTRUCTION AND SEQUENCING:
Each sample took 3 μg total RNA as the starting material to build the circRNA library. A Ribo-Zero Gold kit (human/mouse/rat) was used to remove rRNA from samples and fragment them into short fragments by a fragmentation buffer. The segmented mRNA was used as the template. Random hexamers synthesized the first strand of cDNA, and the second strand of cDNA was synthesized by the buffer, dNTPs (dUTP), RNase H, and DNA polymerase I. The library was purified with a QiaQuick PCR kit, eluted with EB buffer, repaired by the end, added to the base A and sequenced connector, and then amplified by PCR to complete the library preparation. In addition, Qubit 3.0 was used for initial quantification, Agilent 2100 was used for insert size detection of the library, and bio-rad CFX 96 was used to accurately quantify the effective library concentration (effective concentration of the library >10 nM). Lastly, after qualified library detection, the Illumina high-throughput sequencing platform (HiSeq/MiSeq) was used for sequencing. Finally, circRNAs with fold change >2 and
ANALYSIS AND SCREENING OF SEQUENCING RESULTS:
The data were analyzed using CIRI [13] software, compared with circRNAs in the known database (GEO, ENCODE, and EMBL-EBI) using CIRCpedia v2 [14] software, and statistically analyzed. Finally, genes with fold change >2 and P<0.05 were calculated for further differential expression analysis.
ESTABLISHMENT OF CERNA NETWORK:
The differentially expressed circRNAs were sorted according to the different multiples from large to small; the top 5 upregulated and 5 downregulated circRNAs were selected to construct the ceRNA network. Additionally, Circatlas 2.0 [15] software predicted the downstream miRNA combined with circRNAs. Next, Miranda [16] and miRDB [17] software were used to predict downstream binding mRNAs of miRNA. Finally, the mRNA with a target score >80 in the prediction results was compared with the mRNA differentially expressed in our sequencing results to obtain the intersection. The 5 upregulated and 5 downregulated circRNAs, predicted downstream miRNAs, and differentially expressed mRNAs were summarized. The ceRNA network regulating the osteogenic differentiation of rBMSCs was constructed using Cytoscape software.
FUNCTION PREDICTION ANALYSIS OF DIFFERENTIALLY EXPRESSED CIRCRNA AND MRNA:
The GO (
QRT-PCR:
Total RNA was reverse transcribed according to the instructions of the Takara reverse transcription kit. The preparation of the reaction solution was carried out on the ice in a 20-μL reaction system. 1) According to the measured RNA concentration, the formula of v=m/ρ formula was used to calculate the RNA amount of each sample to be added, then gDNA eraser 1 μL and 5×g DNA eraser buffer 2 μL were added. Next, enzyme-free water was added to make the total volume constant as 10 μL, with a reaction at 42°C for 2 min. 2) The second reaction liquid included PrimeScript RT enzyme mix I 1 μL, 5×PrimeScript buffer 24 μL, RT primer mix 1 μL, and RNase free dH2O 4 μL. The above 1) and 2) reaction liquids were mixed, and the program was set to a constant of 37°C, 15 min→85°C, and 5 s→4°C. The obtained cDNA was stored in a refrigerator at −80°C.
The ABI prism 7500 sequence detection system (Applied Biosystems) and power SYBR Green Master Mix (Qiagen, Germany) were used for qRT-PCR. First, the reaction solution was prepared on ice in a 20-μL system. The preparation methods were 2×SYBR Green PCR master mix 10 μL, upstream primer A+1 μL, downstream primer B+1 μL, Rnase-free water 6 μL, and cDNA 2 μL. The reaction conditions were set as 95°C, 2 min→95°C, 5 s→57°C, 30 s, and 40 cycles. The primer sequences are shown in Table 1. Next, the target gene expression was calculated using formula 2−ΔΔCT. Finally, the experiment was repeated 3 times.
STATISTICAL ANALYSIS:
All results were presented as mean±SD of at least 3 independent trials. SPSS 24.0 software (IBM, Armonk, NY, USA) analyzed the data by 1-way ANOVA and
Results
CULTURE AND MULTIDIRECTIONAL DIFFERENTIATION OF RBMSCS:
Microscopically, the primary rBMSCs isolated from the bone marrow cavity of rats showed a typical long spindle shape. After 7 days, the cells adhered to the wall and covered the bottom of the bottle (Figure 1A). After 14 days of osteogenic induction of rBMSCs, alizarine red staining was used to stain the cells, and the formation of a large number of red calcium nodules could be observed, which were characterized by multiple red-stained calcified nodules of different sizes or flake red-stained nodules (Figure 1B). After 14 days of adipogenic induction, the rBMSCs were stained with oil red O staining, and a large number of round or oval red fat droplets were observed (Figure 1C). These results showed that the rBMSCs had multidirectional differentiation ability, which was consistent with the characteristics of bone marrow mesenchymal stem cells.
IDENTIFICATION OF RBMSCS SURFACE MARKERS:
Cell surface markers were used to verify rBMSC phenotypes, and flow cytometry and immunofluorescence analysis were used to identify MSC markers in rBMSCs. The results showed that CD105, CD90, and CD29 were positive (over 90%), whereas CD45 was negative (Figure 2). The results were constant with the characteristics of mesenchymal stem cells.
OSTEOGENIC DIFFERENTIATION OF RBMSCS:
Alizarin red staining indicated that rBMSCs cultured in osteogenic medium for 7 days showed more visible red calcified nodules than rBMSCs cultured in the ordinary medium (Figure 3A). Concurrently, the qRT-PCR revealed that the expression of osteogenic markers ALP, Runx2, BMP2, and COL-1 in osteogenic induced rBMSCs also exhibited a consistent and significantly increased trend during osteogenic differentiation (Figure 3B). These results indicated that bone marrow mesenchymal stem cells were undergoing osteogenic differentiation.
STATISTICS OF SEQUENCING RESULTS OF CIRCRNAS:
First, a box and whisper plot was used to show the differences between 6 groups of samples (3 of osteogenic induction group and 3 of normal control group), with results showing little difference in the intensity of the 6 groups (Figure 4A, 4B). The high-throughput sequencing results showed that 2822 circRNAs were detected during the osteogenesis differentiation of rBMSCs, most of which were derived from sense-overlapping circRNAs (2213 in total). Also, 174 circRNAs were derived from the antisense chain, 202 circRNAs were derived from exons, 140 circRNAs were derived from intergenic regions, and 93 circRNAs were derived from introns (Figure 4C). Our sequencing results overlapped 936 circRNAs out of 9261 circRNAs in the 3 RNA-SEQ databases, GEO, ENCODE, and EMBL-EBI (Figure 4D).
DIFFERENTIAL EXPRESSION OF CIRCRNAS AND MRNAS DURING OSTEOGENIC DIFFERENTIATION OF RBMSCS:
The differential transcripts were analyzed and volcano and heat maps were drawn, with the results showing that there were 29 differentially expressed circRNAs: 13 showed an upward trend during osteogenesis, and 16 showed a downward trend during osteogenesis (Figure 5A, 5B). Similar criteria were used to screen for the differentially expressed mRNAs. The sequencing results revealed that 2453 differentially expressed mRNAs were detected during the osteogenic differentiation of rBMSCs, including 1272 upregulated mRNAs and 1181 downregulated mRNAs (Figure 5C, 5D).
ENRICHMENT ANALYSIS OF DIFFERENTIALLY EXPRESSED CIRCRNAS AND MRNAS:
The differentially expressed circRNAs were analyzed for GO enrichment, and the results showed 223 were differentially enrichment. The results of the first 30 bits were drawn into the GO enrichment analysis diagram. It was mainly concentrated on mechanical stimulation, wound healing, skeletal system development, and cell response to transforming growth factor β stimulus-response, protease binding, and protein binding (Figure 6A). Similarly, GO enrichment analysis screened the differentially expressed mRNAs during osteogenic intramembrane ossification differentiation of rBMSCs. Differential mRNAs enrichment was mainly concentrated in motor tissue-regulation of interleukin-1 β production, positive regulation of cell proliferation, wound healing, negative regulation of gene expression, initiation of DNA replication, protein hybridization activity, protein homopolymerization activity, nucleotide-binding, and structural components of extracellular matrix (Figure 6B).
Additionally, the pathway analysis of the differentiated circRNAs genes was conducted using the KEGG database. The results indicated that 36 differentially enriched KEGG pathways were detected. Similarly, a histogram was drawn for the top 20 KEGG pathways, which mainly included rno04974: protein digestion and absorption; rno04151: PI3K-Akt; rno04933: AGE-RAGE signaling pathway in diabetes complications; rno04512: ECM-receptor interaction; and rno04926: relaxin signaling pathway (Figure 6C). The enrichment analysis of the KEGG public database sorted the pathway of differentially expressed mRNAs, including Rno04218: cell senescence; Rno04151: PI3K-Akt signaling pathway; Rno04110: cell cycle; Rno03030: DNA replication; Rno00480: glutathione metabolism; and Rno00100: steroid biosynthesis (Figure 6D).
CIRCRNA-MIRNA-MRNA REGULATORY NETWORK:
According to the differential multiples, the top 5 upregulated circRNAs and the top 5 downregulated circRNAs in the sequencing results were selected. Since circRNAs contain multiple miRNA binding sites, Miranda and Circatlas 2.0 were used to predict the miRNA bound to these 10 circRNAs. Results showed that 19 miRNAs interacted with 5 upregulated circRNAs and 13 miRNAs interacted with 5 downregulated circRNAs. Miranda and miRDB predicted downstream mRNAs that bind to miRNA and combine with the differentially expressed mRNAs in our sequencing results. The results showed that 13 miRNAs interacting with 5 downregulated circRNAs could bind to 120 downstream mRNAs and 19 miRNAs interacting with 5 upregulated circRNAs could bind to 230 downstream mRNAs. Finally, Cytoscape software was used to construct the circRNA-miRNA-mRNA target interaction network diagram (Figure 7A, 7B).
PCR VALIDATION SEQUENCING RESULTS:
We screened the differentially expressed circRNAs according to their multiple differences to ensure the accuracy and authenticity of the experiment. Five upregulated circRNAs and 5 downregulated circRNAs with the highest multiple difference in the sequencing results were selected for qRT-PCR verification. Also, the results of PCR were consistent with the high-throughput sequencing results. The expression levels of 5 upregulated circRNAs in the osteogenic induction group were significantly increased, compared with the control group. Additionally, the value-added trend of circRNA_1809 was the most visible, almost 3 times that of the control group. The expression of 5 downregulated circRNAs in the osteogenic induction group was also lower than that in the control group; however, the reduction was not as evident as that in the upregulated group, especially circRNAs_2418, which had the largest decline, almost doubled (Figure 8A).
The differentially expressed mRNAs from the high-throughput sequencing results were compared and analyzed with biogenic prediction results, and the common mRNAs of the 2 datasets were retained. Finally, 10 mRNAs related to osteogenesis were screened for PCR verification according to the multiples of their expression changes during the osteogenic differentiation of rBMSCs, which were also 5 upregulated and 5 downregulated. Additionally, the results showed that the expression levels of 5 upregulated mRNAs, Smad1, BMP-2, Sgk1, Fzd4, and Kitlg, significantly increased during osteogenesis, whereas the expression levels of 5 downregulated mRNAs, Wnt5b, Plk4, Cdk1, Klf12, and COL1A2, were considerably decreased, consistent with our sequencing results (Figure 8B).
Discussion
circRNAs are a member of the family of non-coding RNAs owing to the absence of a coding function. They were initially thought to be abnormal RNA splicing products with no function [18]. However, with the recent development of circRNAs research, scientists have found that compared with traditional linear RNA, circRNAs without a 5’ terminal cap and 3’ terminal poly (a) tail possess a more stable closed-loop structure, enabling circRNAs to exist more stably in organisms. It has currently become the focus of recent research on various diseases [19].
Although circRNAs have been implicated in the occurrence and development of cancer and nervous system and cardiovascular diseases, only a few studies have reported the role of circRNAs in osteogenic differentiation. Previous studies suggested that circRNAs play an essential role in the osteogenesis of PDLSC, MSMSCs, and DPSC. For example, Zheng et al [6] conducted RNA sequencing of PDLSC and reported that 139 circRNAs were differentially expressed in D14, establishing a potential ceRNA network relationship. This result provides clues for bone regeneration of PDLSC. However, the osteogenic effect of circRNAs on rBMSCs has not been investigated. Compared with human bone marrow mesenchymal stem cells, rBMSCs have the multi-directional differentiation potential of bone marrow mesenchymal stem cells dividing into bone, cartilage, nerve, and fat and the advantages of being easier to acquire and cultivate. They are a significant source of seed cells in tissue engineering research [20–22]. Therefore, rBMSCs were used as seed cells in this study. The origin of rBMSCs was identified by immunofluorescence and flow cytometry. qRT-PCR established that 4 osteogenic factors, ALP, RUNX2, BMP-2, and COL-1, closely related to osteogenesis, showed a significant upward trend from day 3 to day 14. Simultaneously, when combined with alizarin red staining detection, we can ensure the completion of cell osteogenesis induction. On the 14th day of culture, rBMSCs from the osteogenic induction group and the control group were collected for the high-throughput sequencing to detect alterations in the total transcription expression profile of rBMSCs during osteogenic induction. The results demonstrated that a total of 29 circRNAs and 2453 mRNAs were differentially expressed during the osteogenic differentiation of rBMSCs. Additionally, 13 circRNAs were significantly upregulated, 16 circRNAs were significantly downregulated, 1272 mRNAs were upregulated, and 1181 mRNAs were downregulated.
Next, we performed GO and KEGG analyses on these differentially expressed circRNAs and mRNAs. The GO enrichment analysis results showed that circRNAs and mRNAs were concentrated in wound healing, skeletal system development, intrameminal ossification, protease-binding, protein-binding, and other items that might be related to cell osteogenesis. Further, we found the PI3K-Akt signaling pathway in the differential expression pathway of circRNAs and mRNAs in the KEGG enrichment analysis. The PI3K-Akt signaling pathway is an important pathway involved in regulating cell proliferation and differentiation. Many experiments have proven that the PI3K-Akt signaling pathway is critical in activating osteogenic differentiation [23,24]. For example, Gu et al [25] believe that weakening the PI3K-Akt signaling pathway can inhibit the proliferation of osteoblasts and increase the differentiation of osteoblasts. Moreover, some researchers have noticed the connection between small RNA molecules and the PI3K Akt signaling pathway. For example, Yang et al [26] found that miRNA-21 can activate the PI3K-Akt signaling pathway to promote BMSCs migration and osteogenic differentiation by regulating P-Akt and HIF-1α in vitro. Also, Zheng et al [27] discovered that the overexpression of miRNA-181a would inhibit the expression of the PTEN gene, consequently affecting the transmission of the PI3K-Akt signaling pathway. The above results allow us to speculate that circRNAs may regulate the osteogenic differentiation of rBMSCs, providing clues for the next stage of experiments.
Current studies reveal that circRNAs are closely associated with the growth and development of organisms, including the occurrence and development of diseases; however, their biological functions remain largely not elucidated. Currently, the highly recognized circRNA biological function research mainly focuses on the miRNA molecular sponge, regulating protein binding, regulating gene transcription, and coding function. The circRNAs contain miRNA binding sites to act as an miRNA sponge and regulate downstream gene expression through miRNAs. Considered the most crucial regulation mode of circRNAs, the competitive mechanism involved in regulating gene expression by circRNAs is called ceRNA. Additionally, miRNAs are a member of non-coding RNA. Numerous evidence reports that miRNAs are involved in the regulation mechanism of cell osteogenesis. Some studies proved that miR-26a plays a negative role in the osteogenic differentiation of BMSC and ADSC, while the overexpression of miR-26a weakens the osteogenic ability of BMSCs [28]. Interestingly, owing to the gradual increase in research on circRNAs, the competitive mechanism between circRNAs and miRNAs is being progressively unveiled. Evidence exists that shows hsa_circ_0068871 acts as a molecular sponge of mIR-181a-5p. First, when hsa_circ_0068871 is highly expressed, it competitively binds the target gene FGFR3 with mIR-181a-5p, thus eliminating the inhibitory effect of miR-181a-5p on the target gene and further activating the STAT3 signaling pathway, an essential pathway in promoting the development of bladder cancer [29]. Ji et al [30] studied the relationship between circRNAs and osteoblast differentiation of DPSCs, reporting that hsa_circ_0026827 promotes osteoblast differentiation of DPSCs through mIR-188-3p targeting the Beclin1 and RUNX1 signaling pathways. It also suggests a new treatment for osteoporosis.
Therefore, we speculated that the role of circRNAs in osteogenesis may be related to miRNAs. Firstly, we applied statistical methods to find the 10 circRNAs with the largest changes in molecular expression during osteogenic differentiation. Then, 32 miRNAs that may competitively bind with the 10 circRNAs were obtained using prediction software via continued prediction of the downstream target genes combined with the miRNAs. The predicted results were likened with the differentially expressed mRNAs of high-throughput sequencing, and similar mRNAs were left in the intersection. Additionally, by combining the results of bioinformatics analysis with the results of actual sequencing, we can find the target mRNAs we need more effectively and accurately. The results showed that 13 miRNAs interacting with 5 downregulated circRNAs could bind to 120 downstream mRNAs, and 19 miRNAs interacting with 5 upregulated circRNAs could bind to 230 downstream mRNAs. Finally, Cytoscape software was used to draw the circRNA-miRNA-mRNA interaction network diagram. It was found that the interaction between ceRNA is complicated, the same mRNAs can bind to multiple miRNAs, and the same miRNAs can also bind to multiple circRNAs. Simultaneously, we found some miRNAs in the ceRNA network, which may be related to the osteogenic differentiation of cells. For example, some studies have found that miR-23a-5p is significantly downregulated during osteogenic differentiation. The overexpression of miR-23a-5p inhibits osteogenic differentiation of hBMSCs by targeting MAPK13 [31]. There is also evidence that miR-140-3p regulates osteogenic differentiation of MC3T3-E1 cells as a key factor in the Wnt3a and TGFβ signaling pathways [32]. These data provide us evidence to believe that circRNAs can regulate osteoblastic mRNAs through miRNAs mediation, thus affecting the osteogenic differentiation of rBMSCs.
Finally, we selected the top 5 upregulated circRNAs and 5 downregulated circRNAs in the high-throughput sequencing results for qRT-PCR verification. Among the upregulated circRNAs, circRNA_1809 had the highest degree of upregulation, and the decrease of circRNA_2418 was most apparent. The results of PCR validation were consistent with our sequencing results. Simultaneously, the top 10 osteogenic mRNAs with the highest level of differential expression in the osteogenic differentiation of rBMSCs were screened and were also 5 upregulated and 5 downregulated. The upregulated mRNAs included Smad1, BMP-2, Sgk1, Fzd4, and Kitlg. Many experiments have proven that BMP-2 and Smad1 are critical regulatory factors of osteogenic differentiation. Interestingly, Sgk1 has been reported to promote osteogenic transdifferentiation and calcification of vascular smooth muscle cells under hyperglycemia [33]. Also, Fzd4 promotes proliferation and osteogenic differentiation of hBMSCs through the Wnt/β-catenin pathway [34]. In addition, the verification results of PCR were consistent with the results of our high-throughput sequencing. The expression of these mRNAs significantly increased during the osteogenic differentiation of rBMSCs. Also, Kitlg showed the largest increase, and we found that Kitlg could bind with many miRNAs in the regulatory network of the ceRNA we constructed, such as miR-742-3p, miR-452-5p, miR-196b-3p, miR-3585-3p, and miR-370-3p. Additionally, Kitkg is also known as a stem cell factor (SCF). It plays a vital role in regulating cell survival and proliferation, hematopoiesis, stem cell maintenance, gametogenesis, mast cell development, and cell migration [35]. The Kitlg/SCF combination can activate PI3K-Akt/MAPK and other signal pathways [36]. The downregulated mRNAs include Wnt5b, Plk4, CDK1, Klf12, and COL1A2. The expression of these mRNAs also significantly decreased during the osteogenic differentiation of rBMSCs. The adipogenic and osteogenic differentiation of BMSCs will be affected by the CDR1as-mIR-7-5p-WNT5b axis [37]. Overexpression of Plk4 was shown to significantly inhibit osteoblastic differentiation of hBMSCs through Wnt/β-catenin and BMP/Smad signaling [38]. Klf12 inhibits the osteogenic differentiation of US-Mscs through the regulation of miR-21 [39]. COL1A2 and CDK1 are closely related to osteogenic hypoplasia and osteoporosis [40,41].
In this study, circRNA_1809 and Kitlg were found to be the most upregulated circRNA and mRNA during osteogenic differentiation in rBMSCs. We found that a total of 6 miRNAs in the ceRNA network had competitive binding relationship with circRNA_1809. Further analysis found that miR-370-3p was not only associated with circRNA_1809, but also with downstream Kitlg. Some studies have paid attention to the relationship between miR-370-3p and osteogenic differentiation of bone marrow mesenchymal stem cells, showing the expression of miR-370-3p gradually decreased during the process of osteogenic differentiation in bone marrow mesenchymal stem cells, and the upregulation of miR-370-3p led to the reduction of osteogenic differentiation. Finally, it was proven that LINC00707 could act as a molecular sponge of miR-370-3p and promote the osteogenic differentiation of bone marrow mesenchymal stem cells by upregulating WNT2B [42]. Therefore, we suspect that circRNA_1809 may be involved in the osteogenic differentiation of rat bone marrow mesenchymal stem cells as an miR-370-3p molecular sponge regulatory target gene Kitlg. However, the inference still needs rigorous verification, which is also the research direction of our subsequent experiments.
Conclusions
Therefore, these results suggest that circRNAs may regulate downstream mRNAs by adsorbing miRNAs, and then participate in the osteogenic differentiation of rBMSCs, with circRNA_1809/miR-370-3p/Kitlg may be one of the important regulatory pathways. These results not only lay a foundation for further understanding of the mechanism of circRNAs regulating osteogenic differentiation of rBMSCs but also provide a new idea for the application of circRNA in the repair of bone defects.
Figures
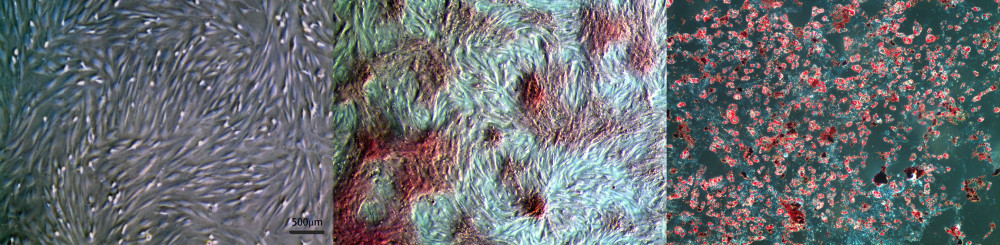 Figure 1. Culture and multidirectional differentiation of bone marrow-derived mesenchymal stem cells (BMSCs). (A) Rat ®BMSCs were isolated and cultured for 7 days (×50). (B) The results of alizarin red staining after 14 days of osteogenic induction of rBMSCs (×50). (C) The results of oil red O staining after 14 days of adipogenic induction of rBMSCs (×50).
Figure 1. Culture and multidirectional differentiation of bone marrow-derived mesenchymal stem cells (BMSCs). (A) Rat ®BMSCs were isolated and cultured for 7 days (×50). (B) The results of alizarin red staining after 14 days of osteogenic induction of rBMSCs (×50). (C) The results of oil red O staining after 14 days of adipogenic induction of rBMSCs (×50). 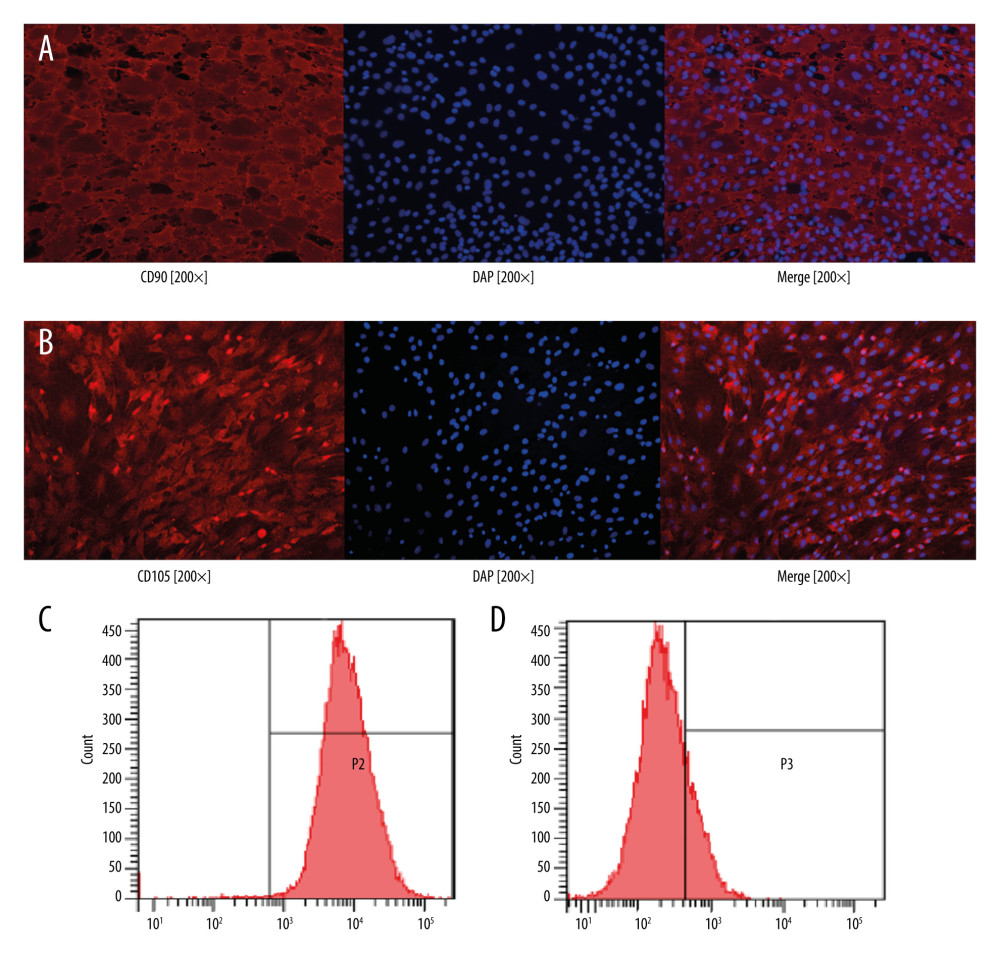 Figure 2. Identification of rat bone marrow-derived mesenchymal stem cell surface markers. (A) Cell surface marker CD90 was positive by immunofluorescence assay. (B) Cell surface marker CD105 was positive by immunofluorescence assay. (C) Cell surface marker CD29 was positive by flow cytometry. (D) Cell surface marker CD45 was negative by flow cytometry.
Figure 2. Identification of rat bone marrow-derived mesenchymal stem cell surface markers. (A) Cell surface marker CD90 was positive by immunofluorescence assay. (B) Cell surface marker CD105 was positive by immunofluorescence assay. (C) Cell surface marker CD29 was positive by flow cytometry. (D) Cell surface marker CD45 was negative by flow cytometry. 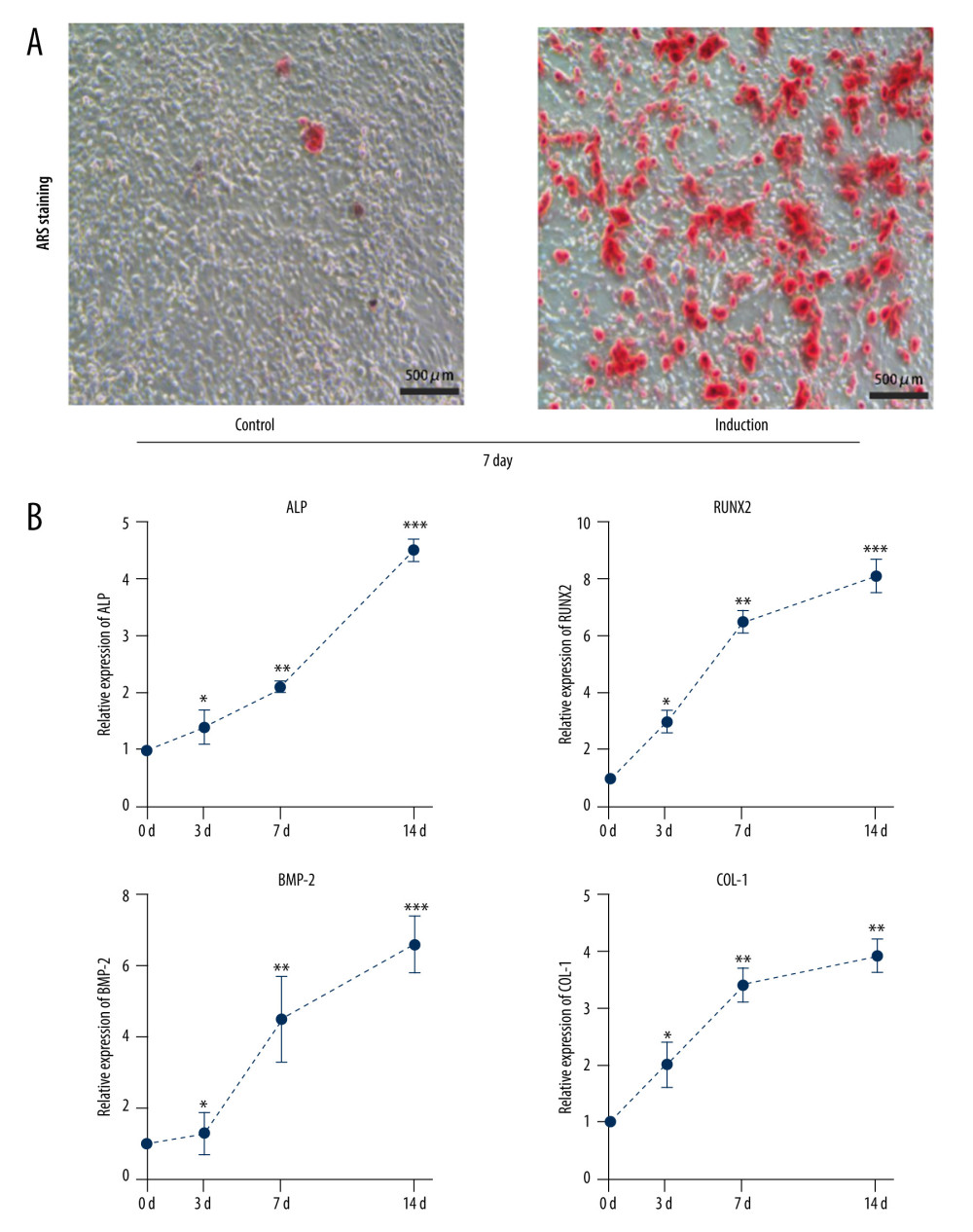 Figure 3. Osteogenic differentiation of rat bone marrow-derived mesenchymal stem cells. (A) Alizarin red staining results of the induced osteogenesis group and the control group. (B) Bone formation markers ALP, BMP2, RUNX2, and COL-1 significantly increased from 3 days to 14 days in the osteogenic induction group detected by qRT-PCR. (GraphPad, Version 8.4.).
Figure 3. Osteogenic differentiation of rat bone marrow-derived mesenchymal stem cells. (A) Alizarin red staining results of the induced osteogenesis group and the control group. (B) Bone formation markers ALP, BMP2, RUNX2, and COL-1 significantly increased from 3 days to 14 days in the osteogenic induction group detected by qRT-PCR. (GraphPad, Version 8.4.). 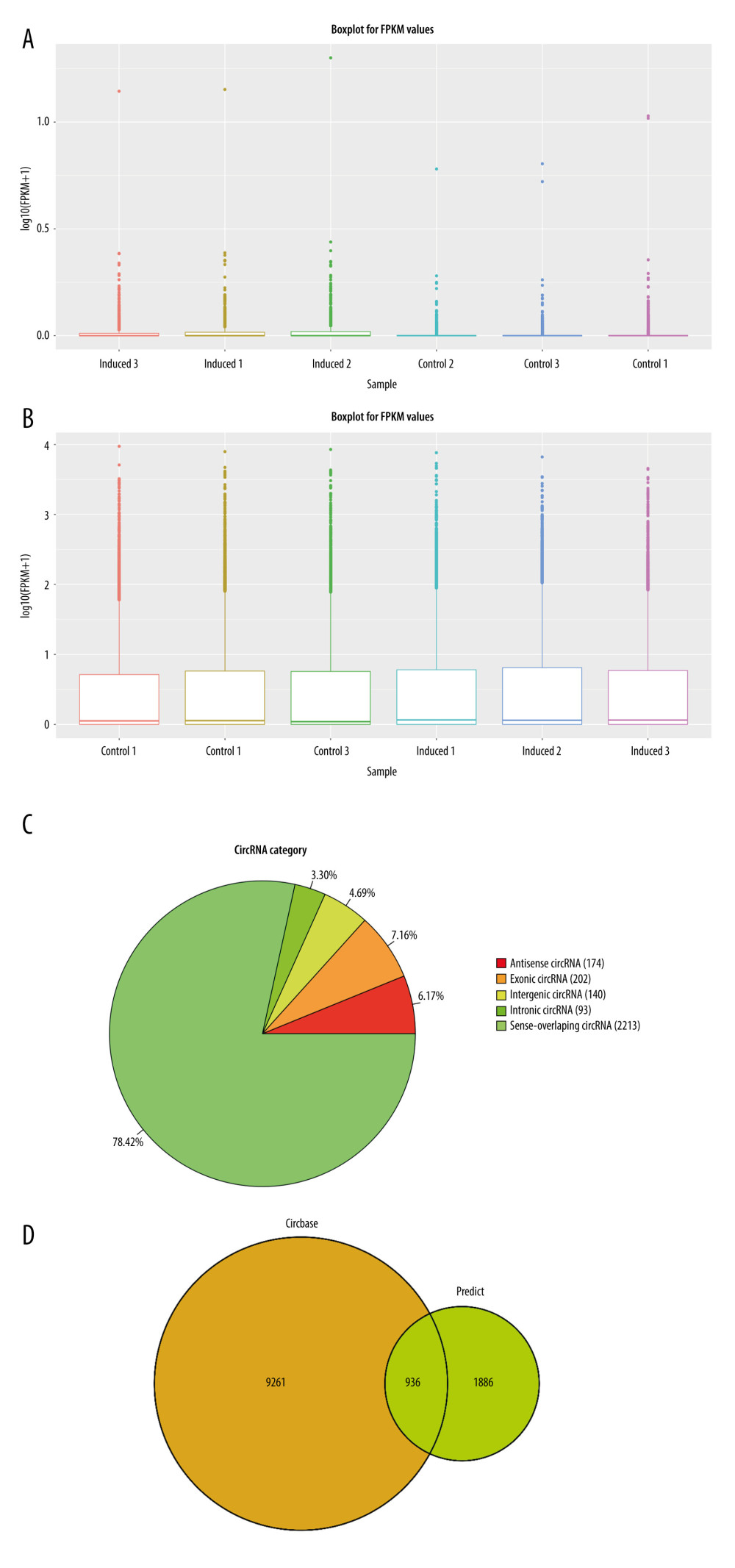 Figure 4. Statistics of sequencing results of circRNAs. (A) Box-whisker plot reflect the signal difference of circRNAs sequencing samples. (B) Box-whisker plot reflect the signal difference of mRNAs sequencing samples. (C) circRNAs gene structure distribution map. (D) The circRNAs identified compared with circRNAs in the known database. (A, C) The heat maps were constructed based on expression values of circRNAs and mRNAs detected by RNA-seq, respectively. The expression values from -2 to 2 were depicted in line with the color scale representing the intensity increased from green to red. Three osteogenic induced groups and 3 control groups were included, and each group included 3 samples. Each column represented 1 sample and each row indicated 1 transcript. (B, D) Filtered transcripts were in gray, transcripts with nonsignificant differences were in blue, and transcripts with significant differences were in red. The log 2 foldchange was displayed on the horizontal axis, and −log 10 (P value) was displayed on the vertical axis.
Figure 4. Statistics of sequencing results of circRNAs. (A) Box-whisker plot reflect the signal difference of circRNAs sequencing samples. (B) Box-whisker plot reflect the signal difference of mRNAs sequencing samples. (C) circRNAs gene structure distribution map. (D) The circRNAs identified compared with circRNAs in the known database. (A, C) The heat maps were constructed based on expression values of circRNAs and mRNAs detected by RNA-seq, respectively. The expression values from -2 to 2 were depicted in line with the color scale representing the intensity increased from green to red. Three osteogenic induced groups and 3 control groups were included, and each group included 3 samples. Each column represented 1 sample and each row indicated 1 transcript. (B, D) Filtered transcripts were in gray, transcripts with nonsignificant differences were in blue, and transcripts with significant differences were in red. The log 2 foldchange was displayed on the horizontal axis, and −log 10 (P value) was displayed on the vertical axis. 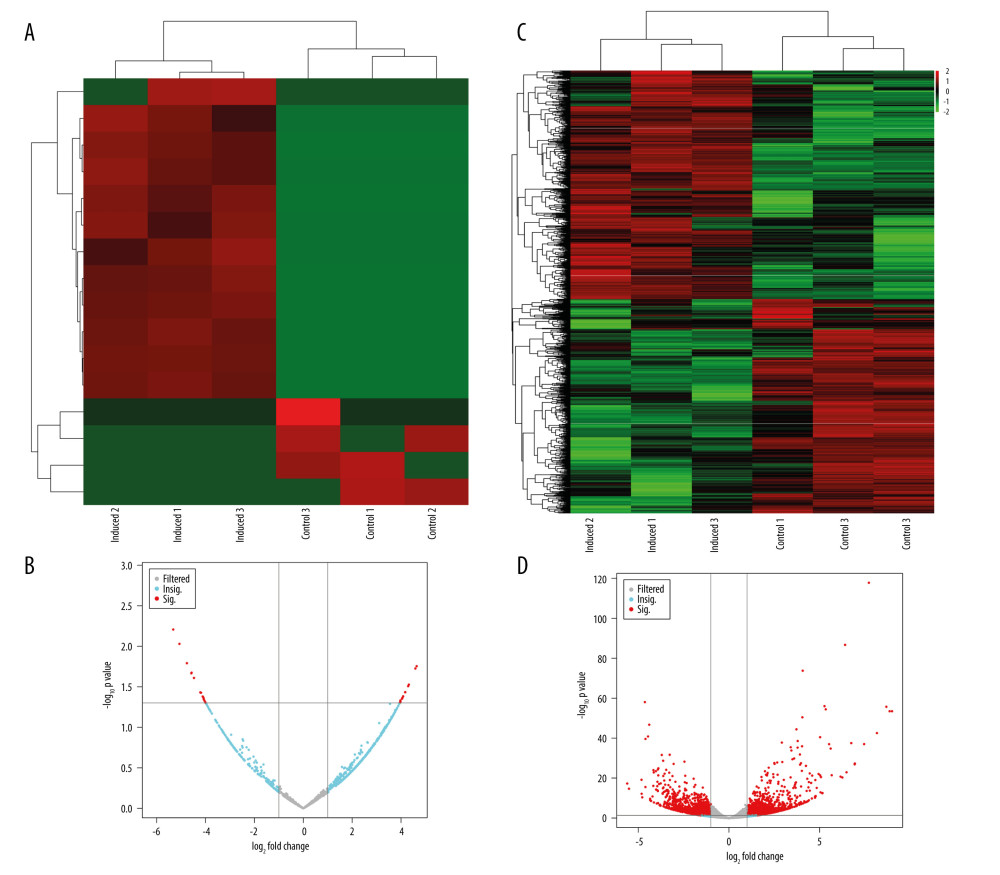 Figure 5. Differential expression of circRNAs and mRNAs. (A) circRNA differential expression heat map. (B) circRNA differential expression volcano map. (C) mRNA differential expression heat map. (D) mRNA differential expression volcano map.
Figure 5. Differential expression of circRNAs and mRNAs. (A) circRNA differential expression heat map. (B) circRNA differential expression volcano map. (C) mRNA differential expression heat map. (D) mRNA differential expression volcano map. 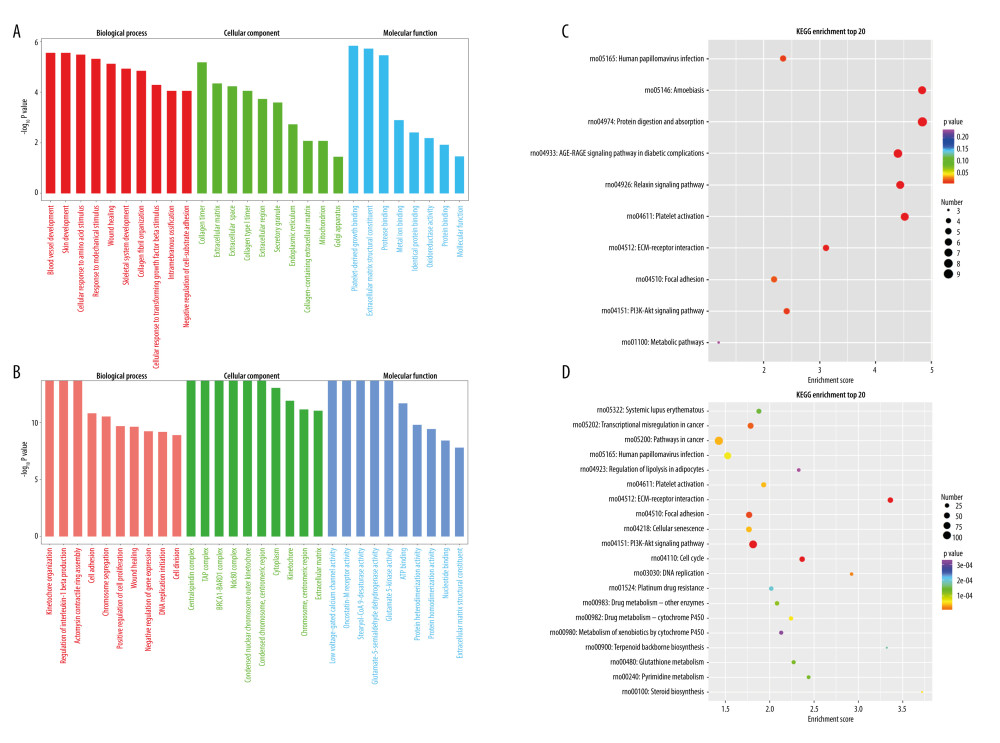 Figure 6. Enrichment analysis of circRNAs and mRNAs. (A) The top 30 GO-enriched entries of differentially expressed circRNAs. (B) The top 30 GO-enriched entries of differentially expressed mRNAs. (C) The top 20 KEGG enrichment pathways of differentially expressed circRNAs. (D) The top 20 KEGG enrichment pathways of differentially expressed mRNAs.
Figure 6. Enrichment analysis of circRNAs and mRNAs. (A) The top 30 GO-enriched entries of differentially expressed circRNAs. (B) The top 30 GO-enriched entries of differentially expressed mRNAs. (C) The top 20 KEGG enrichment pathways of differentially expressed circRNAs. (D) The top 20 KEGG enrichment pathways of differentially expressed mRNAs. 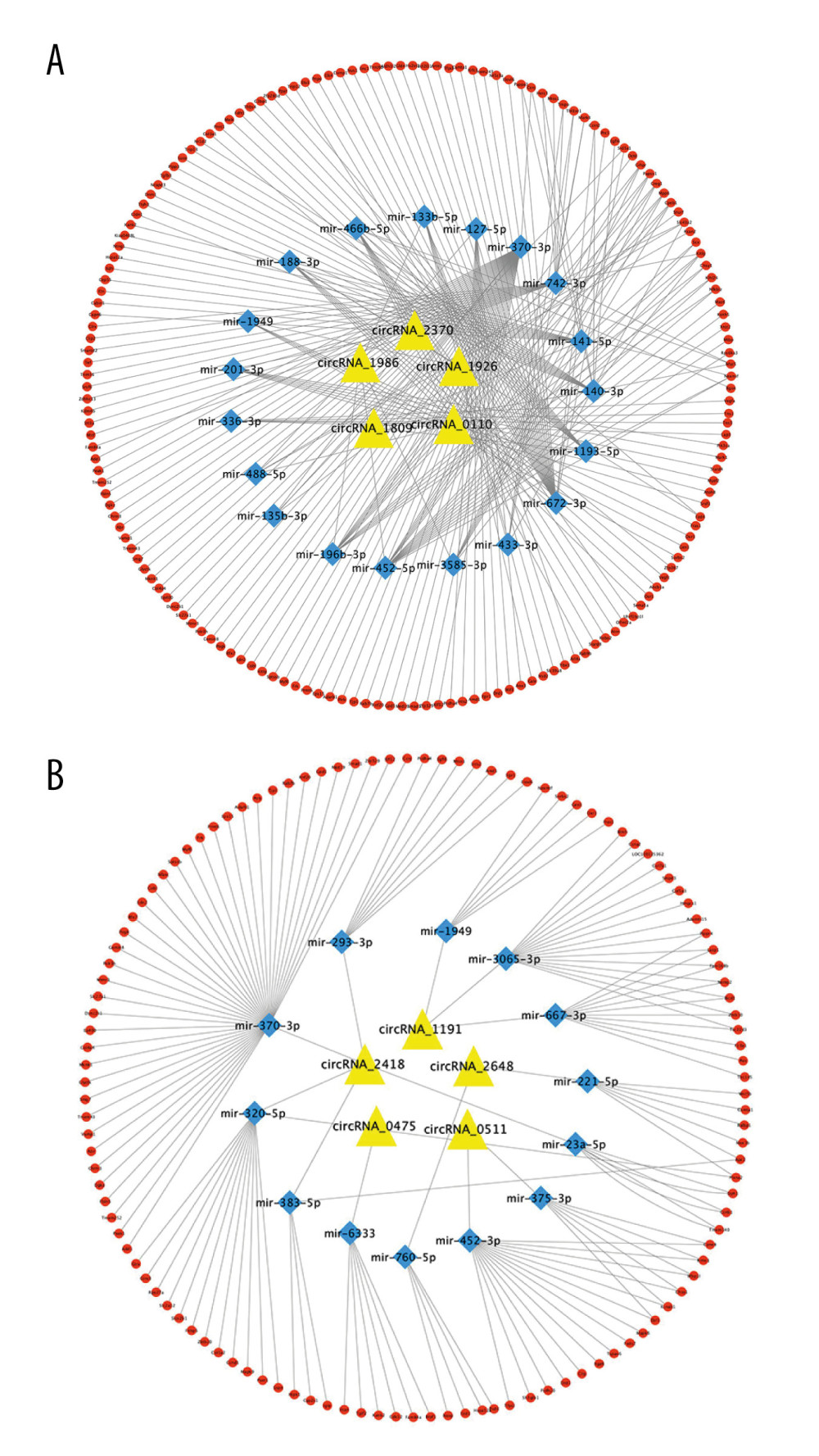 Figure 7. The circRNA-miRNA-mRNA regulatory network. (A) ceRNA network of the first 5 upregulated circRNAs. (B) ceRNA network of the first 5 downregulated circRNAs. (Cytoscape,version 3.6.1.). The statistical results were presented as mean±standard deviation; * P<0.05, ** P<0.01, *** P<0.001.
Figure 7. The circRNA-miRNA-mRNA regulatory network. (A) ceRNA network of the first 5 upregulated circRNAs. (B) ceRNA network of the first 5 downregulated circRNAs. (Cytoscape,version 3.6.1.). The statistical results were presented as mean±standard deviation; * P<0.05, ** P<0.01, *** P<0.001. 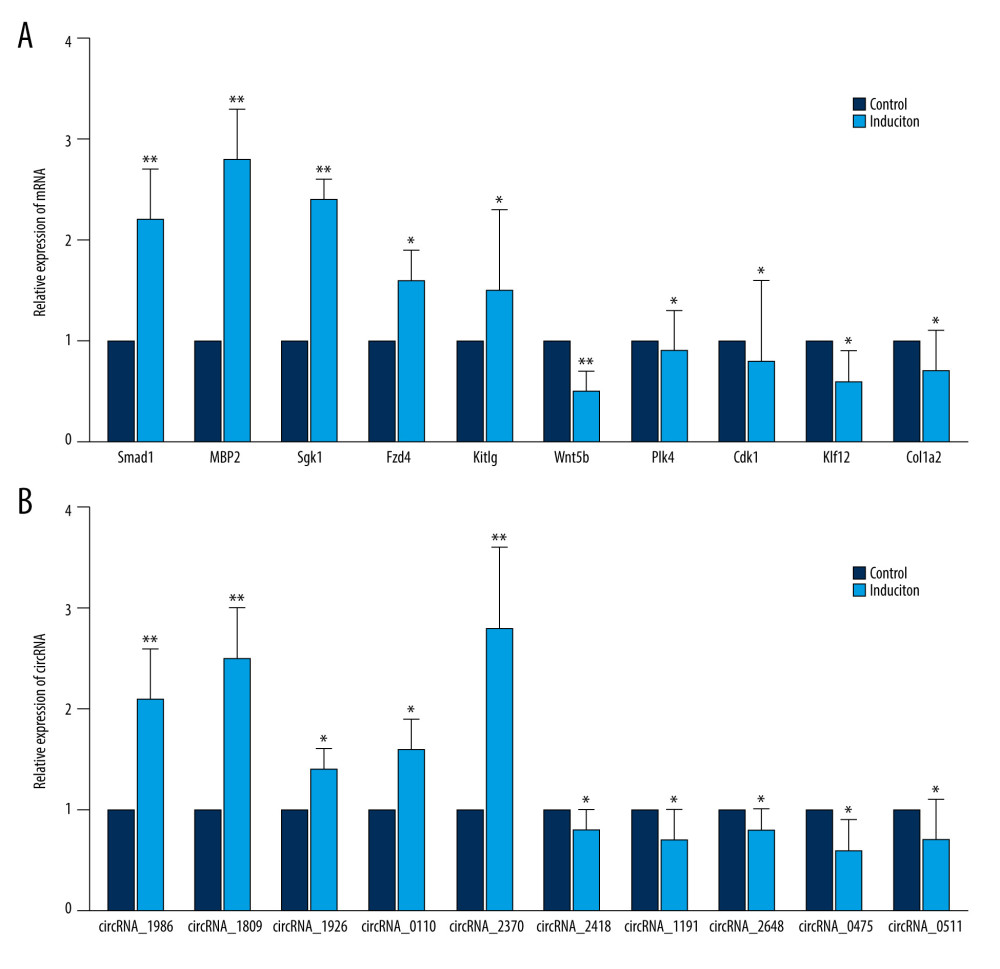 Figure 8. PCR validation sequencing results. (A) qRT-PCR verified the top 5 upregulated and 5 downregulated circRNAs in the sequencing results. (B) qRT-PCR verified the top 5 upregulated and 5 downregulated mRNAs in the sequencing results. (GraphPad, Version 8.4.).
Figure 8. PCR validation sequencing results. (A) qRT-PCR verified the top 5 upregulated and 5 downregulated circRNAs in the sequencing results. (B) qRT-PCR verified the top 5 upregulated and 5 downregulated mRNAs in the sequencing results. (GraphPad, Version 8.4.). 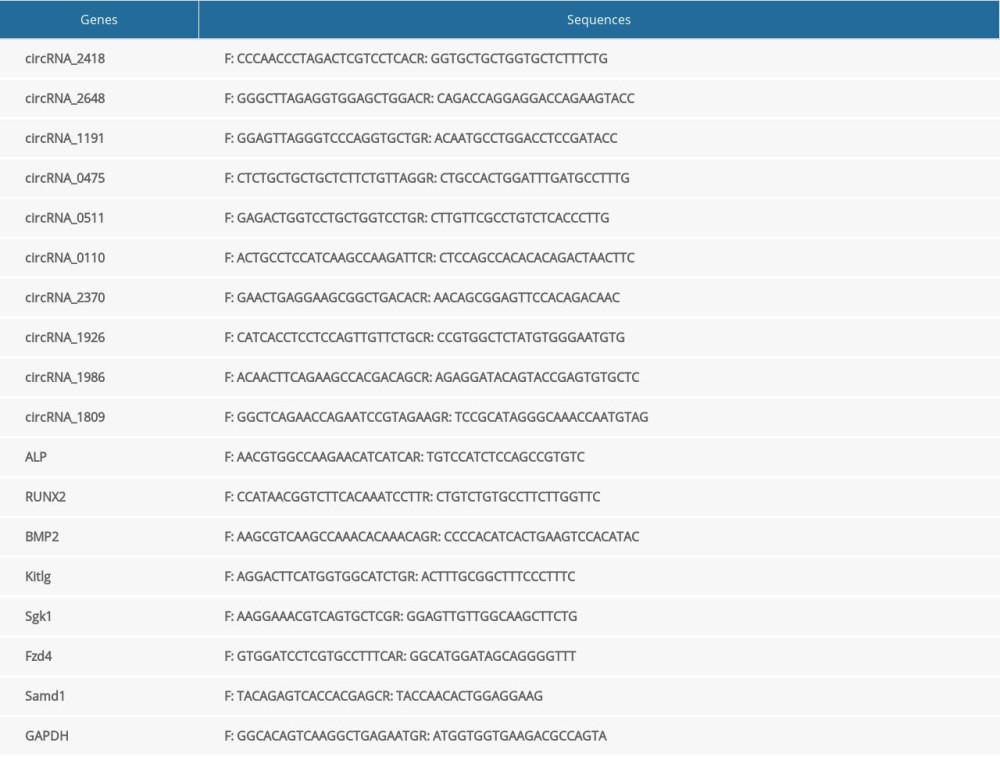
References
1. Zeng JH, Liu SW, Xiong L, Scaffolds for the repair of bone defects in clinical studies: A systematic review: J Orthop Surg Res, 2018; 13(1); 33
2. Tamaddon M, Burrows M, Ferreira SA, Monomeric, porous type II collagen scaffolds promote chondrogenic differentiation of human bone marrow mesenchymal stem cells in vitro: Sci Rep, 2017; 7; 43519
3. Matic I, Antunovic M, Brkic S, Expression of OCT-4 and SOX-2 in bone marrow-derived human mesenchymal stem cells during osteogenic differentiation: Open Access Maced J Med Sci, 2016; 4(1); 9-16
4. Qin Y, Wang L, Gao Z, Bone marrow stromal/stem cell-derived extracellular vesicles regulate osteoblast activity and differentiation in vitro and promote bone regeneration in vivo: Sci Rep, 2016; 6; 21961
5. Yusop N, Battersby P, Alraies A, Isolation and characterisation of mesenchymal stem cells from rat bone marrow and the endosteal niche: A comparative study: Stem Cells Int, 2018; 2018; 6869128
6. Zheng Y, Li X, Huang Y, The circular RNA landscape of periodontal ligament stem cells during osteogenesis: J Periodontol, 2017; 88(9); 906-14
7. Gao Y, Wang J, Zhao F, CIRI: An efficient and unbiased algorithm for de novo circular RNA identification: Genome Biol, 2015; 16(1); 4
8. Zhang M, Jia L, Zheng Y, circRNA expression profiles in human bone marrow stem cells undergoing osteoblast differentiation: Stem Cell Rev Rep, 2019; 15(1); 126-38
9. Hansen TB, Jensen TI, Clausen BH, Natural RNA circles function as efficient microRNA sponges: Nature, 2013; 495(7441); 384-88
10. Lu Q, Liu T, Feng H, Circular RNA circSLC8A1 acts as a sponge of miR-130b/miR-494 in suppressing bladder cancer progression via regulating PTEN: Mol Cancer, 2019; 18(1); 111
11. Du Y, Li J, Hou Y, Alteration of circular RNA expression in rat dental follicle cells during osteogenic differentiation: J Cell Biochem, 2019; 120(8); 13289-301
12. Gu X, Li M, Jin Y, Identification and integrated analysis of differentially expressed lncRNAs and circRNAs reveal the potential ceRNA networks during PDLSC osteogenic differentiation: BMC Genet, 2017; 18(1); 100
13. Gao Y, Wang J, Zhao F, CIRI: An efficient and unbiased algorithm for de novo circular RNA identification: Genome Biol, 2015; 16; 4
14. Dong R, Ma XK, Li GW, CIRCpedia v2: An updated database for comprehensive circular RNA annotation and expression comparison: GENOM Proteom Bioinf, 2018; 16(4); 226-33
15. Wu W, Ji P, Zhao F, CircAtlas: an integrated resource of one million highly accurate circular RNAs from 1070 vertebrate transcriptomes: Genome Biol, 2020; 21(1); 101
16. Enright AJ, John B, Gaul U, MicroRNA targets in Drosophila: Genome Biol, 2003; 5(1); e363
17. Chen Y, Wang X, miRDB: An online database for prediction of functional microRNA targets: Nucleic Acids Res; 48(1); 127-31
18. Della Bella E, Menzel U, Basoli V, Differential regulation of circRNA, miRNA, and piRNA during early osteogenic and chondrogenic differentiation of human mesenchymal stromal cells: Cells, 2020; 9(2); 398
19. Xu X, Chen Y, Tan B, Circular RNA circ_0011269 sponges miR-122 to regulate RUNX2 expression and promotes osteoporosis progression: J Cell Biochem, 2020 [Online ahead of print]
20. Miao Z, Lu Z, Luo S, Murine and Chinese cobra venom-derived nerve growth factor stimulate chondrogenic differentiation of BMSCs in vitro: A comparative study: Mol Med Rep, 2018; 18(3); 3341-49
21. Ruan H, Xiao R, Jiang X, Biofunctionalized self-assembly of peptide amphiphile induces the differentiation of bone marrow mesenchymal stem cells into neural cells: Mol Cell Biochem, 2019; 450(1–2); 199-207
22. Chen C, Tian Y, Wang J, Testosterone propionate can promote effects of acellular nerve allograft-seeded bone marrow mesenchymal stem cells on repairing canine sciatic nerve: J Tissue Eng Regen Med, 2019; 13(9); 1685-701
23. Liu H, Li X, Lin J, Morroniside promotes the osteogenesis by activating PI3K/Akt/mTOR signaling: Biosci Biotechnol Biochem, 2021; 85(2); 332-39
24. Yu RH, Zhang XY, Xu W, Apolipoprotein D alleviates glucocorticoid-induced osteogenesis suppression in bone marrow mesenchymal stem cells via the PI3K/Akt pathway: J Orthop Surg Res, 2020; 15(1); 307
25. Gu YX, Du J, Si MS, The roles of PI3K/Akt signaling pathway in regulating MC3T3-E1 preosteoblast proliferation and differentiation on SLA and SLActive titanium surfaces: J Biomed Mater Res A, 2013; 101(3); 748-54
26. Yang C, Liu X, Zhao K, miRNA-21 promotes osteogenesis via the PTEN/PI3K/Akt/HIF-1α pathway and enhances bone regeneration in critical size defects: Stem Cell Res Ther, 2019; 10(1); 65
27. Zheng H, Liu J, Tycksen E, MicroRNA-181a/b-1 over-expression enhances osteogenesis by modulating PTEN/PI3K/AKT signaling and mitochondrial metabolism: Bone, 2019; 123; 92-102
28. Su X, Liao L, Shuai Y, MiR-26a functions oppositely in osteogenic differentiation of BMSCs and ADSCs depending on distinct activation and roles of Wnt and BMP signaling pathway: Cell Death Dis, 2015; 6; e1851
29. Mao W, Huang X, Wang L, Circular RNA hsa_circ_0068871 regulates FGFR3 expression and activates STAT3 by targeting miR-181a-5p to promote bladder cancer progression: J Exp Clin Cancer Res, 2019; 38(1); 169
30. Ji F, Zhu L, Pan J, hsa_circ_0026827 promotes osteoblast differentiation of human dental pulp stem cells through the Beclin1 and RUNX1 signaling pathways by sponging miR-188-3p: Front Cell Dev Biol, 2020; 8; 470
31. Ren G, Sun J, Li MM, MicroRNA-23a-5p regulates osteogenic differentiation of human bone marrow-derived mesenchymal stem cells by targeting mitogen-activated protein kinase-13: Mol Med Rep, 2018; 17(3); 4554-60
32. Fushimi S, Nohno T, Nagatsuka H, Involvement of miR-140-3p in Wnt3a and TGFβ3 signaling pathways during osteoblast differentiation in MC3T3-E1 cells: Genes Cells, 2018; 23(7); 517-27
33. Poetsch F, Henze LA, Estepa M, Role of SGK1 in the osteogenic transdifferentiation and calcification of vascular smooth muscle cells promoted by hyperglycemic conditions: Int J Mol Sci, 2020; 21(19); 7207
34. Zhang Z, Zhou H, Sun F, Circ_FBLN1 promotes the proliferation and osteogenic differentiation of human bone marrow-derived mesenchymal stem cells by regulating let-7i-5p/FZD4 axis and Wnt/β-catenin pathway: J Bioenerg Biomembr, 2021; 53(5); 561-72
35. Cao Z, Xie Y, Yu L, Hepatocyte growth factor (HGF) and stem cell factor (SCF) maintained the stemness of human bone marrow mesenchymal stem cells (hBMSCs) during long-term expansion by preserving mitochondrial function via the PI3K/AKT, ERK1/2, and STAT3 signaling pathways: Stem Cell Res Ther, 2020; 11(1); 329
36. Thuwanut P, Comizzoli P, Wildt DE, Stem cell factor promotes in vitro ovarian follicle development in the domestic cat by upregulating c-kit mRNA expression and stimulating the phosphatidylinositol 3-kinase/AKT pathway: Reprod Fertil Dev, 2017; 29(7); 1356-68
37. Chen G, Wang Q, Li Z, Circular RNA CDR1as promotes adipogenic and suppresses osteogenic differentiation of BMSCs in steroid-induced osteonecrosis of the femoral head: Bone, 2020; 133; 115258
38. Zhou B, Peng K, Wang G, Polo Like Kinase 4 (PLK4) impairs human bone marrow mesenchymal stem cell (BMSC) viability and osteogenic differentiation: Biochem Biophys Res Commun, 2021; 549; 221-28
39. Zhao G, Luo WD, Yuan Y, LINC02381, a sponge of miR-21, weakens osteogenic differentiation of hUC-MSCs through KLF12-mediated Wnt4 transcriptional repression: J Bone Miner Metab, 2022; 40(1); 66-80
40. Kim JH, Nam SW, Kim BW, Astaxanthin improves the proliferative capacity as well as the osteogenic and adipogenic differentiation potential in neural stem cells: Food Chem Toxicol, 2010; 48(6); 1741-45
41. Yang L, Liu B, Dong X, Clinical severity prediction in children with osteogenesis imperfecta caused by COL1A1/2 defects: Osteoporos Int, 2022; 10; 1007
42. Jia B, Wang Z, Sun X, Long noncoding RNA LINC00707 sponges miR-370-3p to promote osteogenesis of human bone marrow-derived mesenchymal stem cells through upregulating WNT2B: Stem Cell Res Ther, 2019; 10(1); 67
Figures
Figure 1. Culture and multidirectional differentiation of bone marrow-derived mesenchymal stem cells (BMSCs). (A) Rat ®BMSCs were isolated and cultured for 7 days (×50). (B) The results of alizarin red staining after 14 days of osteogenic induction of rBMSCs (×50). (C) The results of oil red O staining after 14 days of adipogenic induction of rBMSCs (×50).Figure 2. Identification of rat bone marrow-derived mesenchymal stem cell surface markers. (A) Cell surface marker CD90 was positive by immunofluorescence assay. (B) Cell surface marker CD105 was positive by immunofluorescence assay. (C) Cell surface marker CD29 was positive by flow cytometry. (D) Cell surface marker CD45 was negative by flow cytometry.Figure 3. Osteogenic differentiation of rat bone marrow-derived mesenchymal stem cells. (A) Alizarin red staining results of the induced osteogenesis group and the control group. (B) Bone formation markers ALP, BMP2, RUNX2, and COL-1 significantly increased from 3 days to 14 days in the osteogenic induction group detected by qRT-PCR. (GraphPad, Version 8.4.).Figure 4. Statistics of sequencing results of circRNAs. (A) Box-whisker plot reflect the signal difference of circRNAs sequencing samples. (B) Box-whisker plot reflect the signal difference of mRNAs sequencing samples. (C) circRNAs gene structure distribution map. (D) The circRNAs identified compared with circRNAs in the known database. (A, C) The heat maps were constructed based on expression values of circRNAs and mRNAs detected by RNA-seq, respectively. The expression values from -2 to 2 were depicted in line with the color scale representing the intensity increased from green to red. Three osteogenic induced groups and 3 control groups were included, and each group included 3 samples. Each column represented 1 sample and each row indicated 1 transcript. (B, D) Filtered transcripts were in gray, transcripts with nonsignificant differences were in blue, and transcripts with significant differences were in red. The log 2 foldchange was displayed on the horizontal axis, and −log 10 (P value) was displayed on the vertical axis.Figure 5. Differential expression of circRNAs and mRNAs. (A) circRNA differential expression heat map. (B) circRNA differential expression volcano map. (C) mRNA differential expression heat map. (D) mRNA differential expression volcano map.Figure 6. Enrichment analysis of circRNAs and mRNAs. (A) The top 30 GO-enriched entries of differentially expressed circRNAs. (B) The top 30 GO-enriched entries of differentially expressed mRNAs. (C) The top 20 KEGG enrichment pathways of differentially expressed circRNAs. (D) The top 20 KEGG enrichment pathways of differentially expressed mRNAs.Figure 7. The circRNA-miRNA-mRNA regulatory network. (A) ceRNA network of the first 5 upregulated circRNAs. (B) ceRNA network of the first 5 downregulated circRNAs. (Cytoscape,version 3.6.1.). The statistical results were presented as mean±standard deviation; * P<0.05, ** P<0.01, *** P<0.001.Figure 8. PCR validation sequencing results. (A) qRT-PCR verified the top 5 upregulated and 5 downregulated circRNAs in the sequencing results. (B) qRT-PCR verified the top 5 upregulated and 5 downregulated mRNAs in the sequencing results. (GraphPad, Version 8.4.). In Press
07 Mar 2024 : Clinical Research
Knowledge of and Attitudes Toward Clinical Trials: A Questionnaire-Based Study of 179 Male Third- and Fourt...Med Sci Monit In Press; DOI: 10.12659/MSM.943468
08 Mar 2024 : Animal Research
Modification of Experimental Model of Necrotizing Enterocolitis (NEC) in Rat Pups by Single Exposure to Hyp...Med Sci Monit In Press; DOI: 10.12659/MSM.943443
18 Apr 2024 : Clinical Research
Comparative Analysis of Open and Closed Sphincterotomy for the Treatment of Chronic Anal Fissure: Safety an...Med Sci Monit In Press; DOI: 10.12659/MSM.944127
08 Mar 2024 : Laboratory Research
Evaluation of Retentive Strength of 50 Endodontically-Treated Single-Rooted Mandibular Second Premolars Res...Med Sci Monit In Press; DOI: 10.12659/MSM.944110
Most Viewed Current Articles
17 Jan 2024 : Review article
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
14 Dec 2022 : Clinical Research
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387
01 Jan 2022 : Editorial
Editorial: Current Status of Oral Antiviral Drug Treatments for SARS-CoV-2 Infection in Non-Hospitalized Pa...DOI :10.12659/MSM.935952
Med Sci Monit 2022; 28:e935952